Probiotics have been getting a lot of attention recently. These bacteria, which you can consume from fermented foods, yogurt or even pills, are linked to a number of health and wellness benefits, including reducing gastrointestinal distress, urinary tract infections and eczema. But can they improve your mood, too?
Behavior and mental health are complicated. But the short answer, according to my team’s recently published research, is likely yes.
The beneficial bacteria in probiotics become part of a community of other microscopic organisms living in your digestive system called the gut microbiome. Your gut microbiome contains trillions of a diverse range of bacteria, fungi and viruses.
Hundreds of species of bacteria are native to the intestinal tract. Each species can be broken down into hundreds of strains that can also be dramatically different from each other in their metabolism, byproducts and environmental preferences.
This bacterial diversity is why not all probiotics are built the same. Many research groups have shown that specific strains of Lactobacillus have mood-enhancing effects.
But these effects seem to happen only with the right mix of bacteria in the right conditions. For example, a probiotic that can reduce symptoms of stress in someone who is worried about their calculus final may not work in someone with symptoms of post-traumatic stress disorder.
The probiotics in your yogurt may play a role in boosting mood.
Studying mood in mice
In my work as a neuroscientist, I study how the gut influences the brain. My team and I recently conducted experiments in mice that support the idea that gut microbiota play a role in regulating stress.
So how do you measure the mood of mice?
First, we needed to understand how stressed mice behave. So we placed them under short periods of stress: They are restrained for two hours each day, given enough room to move around but not enough to groom or stand up. We envision this as the same type of stress people experience when they’re confined to a car or cubical for hours at a time.
Stressed mice soon exhibited depression- and anxiety-like behaviors, which we measured by monitoring how much time they spent hiding when placed in a new environment or how quickly they try to right themselves when flipped upside down.
While it isn’t surprising that stressed mice hide longer and are slower to right themselves, the power of their poop to change their behavior was.
To see if stressed behavior could be transferred through the microbiome, we used another group of mice that were entirely clean. These mice were free from any bacteria, fungi or viruses and lived in a rubber bubble. They essentially had no microbiome at all.
We exposed them to poop from either stressed mice or normal mice by sprinkling soiled bedding in their enclosures. Microbes from the donor mice started to populate the gut microbiomes of the clean mice.
Within a few weeks, the clean mice exposed to poop from stressed mice started to develop stress- and anxiety-like behavior, even though nothing else had changed. Meanwhile, clean mice exposed to poop from normal mice had no differences in their behavior. This finding suggests that the microbes in poop changed the mice’s behavior.
Which bacteria affect mood?
The results of our experiments led us back to our original question: Which bacteria can change your mood?
We started by comparing the microbes in the poop of stressed and normal mice. In our analysis, we found that a group of bacteria called Lactobacillus was greatly reduced in the stressed mice. Research has linked this group of bacteria to stress reduction before. However, Lactobacillus contains over 170 different species and even more strains.
Currently, the probiotic supplements available to patients are unregulated and often untested. In order to reliably get the most effective strains to patients, they need to be properly tested. So we had to come up with a way to test how different strains affect anxious behavior.
Lactobacillus are a diverse range of bacteria that can provide potential health benefits in people.
Instead of tackling this colossal task alone, we created a method that other microbiome scientists can also use to look at this group of bacteria as systematically as possible.
To recreate the same experimental conditions for each species of microbe, we created a group of mice with only six species of bacteria in their microbiome, the bare minimum needed for normal and healthy development, which did not include Lactobacillus. This way, we could add individual strains of Lactobacillus back into the mice’s gut microbiome and observe the effects of each strain on their behavior and biology.
We’ve tested two strains so far: Lactobacillus intestinalis ASF360 and Lactobacillus murinus ASF361. Mice with these two strains of Lactobacillus are more resilient to stress and have quieted neural pathways associated with fear.
What’s next?
Our study on how different strains of Lactobacillus affect mood is just the beginning. We hope that our research will open avenues for other scientists to test different probiotics.
While researchers are reaching a consensus that the bacteria in your digestive tract can influence your mood, and vice versa, there is still a lot of testing to be done in both animals and in people.
Our team is starting to develop ways to systematically test which bacteria may provide the best health outcomes in people and which probiotics are the most effective. In the meantime, give the Lactobacillus in your gut some love through a healthy, probiotics-rich diet.
A team of researchers has discovered a method to activate a bacterial defense system, known as CBASS, to self-destruct and prevent the spread of viruses among bacteria, potentially offering a new way to manage bacterial infections and combat antibiotic resistance. Credit: SciTechDaily.com
Researchers unveil how the self-killing activity of bacteria can be harnessed in the fight against antibiotic resistance.
Scientists at the Icahn School of Medicine at Mount Sinai have identified a new approach to controlling bacterial infections. The findings were described in the February 6 online issue of Nature Structural & Molecular Biology.
The team found a way to turn on a vital bacterial defense mechanism to fight and manage bacterial infections. The defense system, called cyclic oligonucleotide-based antiphage signaling system (CBASS), is a natural mechanism used by certain bacteria to protect themselves from viral attacks. Bacteria self-destruct as a means to prevent the spread of virus to other bacterial cells in the population.
CBASS Defense Mechanism Explored
“We wanted to see how the bacterial self-killing CBASS system is activated and whether it can be leveraged to limit bacterial infections,” says co-senior author Aneel Aggarwal, PhD, Professor of Pharmacological Sciences at Icahn Mount Sinai. “This is a fresh approach to tackling bacterial infections, a significant concern in hospitals and other settings. It’s essential to find new tools for fighting antibiotic resistance. In the war against superbugs, we need to constantly innovate and expand our toolkit to stay ahead of evolving drug resistance.”
Icahn Mount Sinai researchers unveil how the self-killing activity of bacteria can be used in the fight against antibiotic resistance. Above: 3-D structure of CBASS Cap5 protein tetramer (shown in cyan) formed upon binding to the cyclic dinucleotide (shown in orange) to destroy bacteria’s own DNA (model, shown in red). Essential magnesium ions for DNA cleavage are shown in green.
Innovative Strategies Against Superbugs
As part of the experiments, the researchers studied how “Cap5,” or CBASS-associated protein 5, is activated for DNA degradation and how it could be used to control bacterial infections through a combination of structural analysis and various biophysical, biochemical, and cellular assays. Cap5 is a key protein that becomes activated by cyclic nucleotides (small signaling molecules) to destroy the bacterial cell’s own DNA.
“In our study, we started by identifying which of the many cyclic nucleotides could activate the effector Cap5 of the CBASS system,” says co-senior author Olga Rechkoblit, PhD, Assistant Professor of Pharmacological Sciences at Icahn Mount Sinai. “Once we figured that out, we looked closely at the structure of Cap5 when it’s bound to these small signaling molecules. Then, with expert help from Daniela Sciaky, PhD, a researcher at Icahn Mount Sinai, we showed that by adding these special molecules to the bacteria’s environment, these molecules could potentially be used to eliminate the bacteria.”
Overcoming Technical Challenges
The researchers found that determining the structure of Cap5 with cyclic nucleotides posed a technical challenge, requiring expert help from Dale F. Kreitler, PhD, AMX Beamline Scientist at Brookhaven National Laboratory. It was achieved by using micro-focused synchrotron X-ray radiation at the same facility. Micro-focused synchrotron X-ray radiation is a type of X-ray radiation that is not only produced using a specific type of particle accelerator (synchrotron) but is also carefully concentrated or focused on a tiny area for more detailed imaging or analysis.
Future Directions
Next, the researchers will explore how their discoveries apply to other types of bacteria and assess whether their method can be used to manage infections caused by various harmful bacteria
These are bacteria devastated by attack with the toxic arrowhead. Credit: T.E. Wood et al. (2019)
A weapon bacteria use to vanquish their competitors could be copied to create new forms of antibiotics, according to Imperial College London research.
Researchers have uncovered a novel weapon in the arsenal of bacteria that works in a similar way to common antibiotics.
“By further understanding and characterizing the molecular targets of VgrG2b, and how the toxin works, this research would support the design of new antibiotics.” — Dr. Thomas Wood
By discovering the mechanism of this new weapon, the team says new antibiotics could be designed to fill the gap left by rising antibiotic resistance.
Lead author Professor Alain Filloux, from the MRC Center for Molecular Bacteriology and Infection in the Department of Life Sciences at Imperial, said: “Bacteria evolved alongside each other for billions of years and have designed many strategies to fight and kill each other to prevail in the environment.
“This new ‘toxic arrowhead’ is one of the tools developed for this, and finding all the other possible strategies employed for bacterial warfare would help researchers and the pharmaceutical industry replenish the currently dwindling pipeline of antibiotics.”
Fired from a molecular crossbow
The new study, published in Cell Reports, looked at the Type VI Secretion System (T6SS) – a ‘molecular crossbow’ that some bacteria use to launch toxic packages at rival bacteria. Bacteria fight for dominance to control food sources and space, allowing them to thrive in their environment.
“Bacteria evolved alongside each other for billions of years and have designed many strategies to fight and kill each other to prevail in the environment.” — Professor Alain Filloux
The team discovered a new kind of toxic arrowhead used by the bacteria Pseudomonas aeruginosa, called VgrG2b, with the toxin found at the very tip. By analyzing the atomic structure of the toxin, the team showed that it is an enzyme known as a metallopeptidase, which cuts up proteins.
Once fired into a rival bacterium, VgrG2b targets the cell envelope, between the inner and outer bacterial membranes and encompassing the cell wall. Dr. Abderrahman Hachani, now at the University of Melbourne’s Doherty Institute, said: “This vital area provides structural integrity to the cell, regulates its growth, and acts as a protective barrier against the external environment.”
VgrG2b prevents the cell from dividing, causing it instead to bulge and eventually explode as the integrity of the cell fails.
Antibiotic mimic
This form of attack is similar to how a group of antibiotics, called beta-lactams, work. These antibiotics include penicillin, and as is the case with many classes of antibiotics, many harmful bacteria are becoming increasingly resistant to them.
Dr. Thomas Wood, now at Harvard Medical School, said: “The impact of VgrG2b on target cells mimics the action of beta-lactam antibiotics. Yet it is clear that its mode of action is different. By further understanding and characterizing the molecular targets of VgrG2b, and how the toxin works, this research would support the design of new antibiotics.”
Sometimes the therapeutic effects of honey surpass even those of medication.
In this series, we will explore the good and bad sweeteners, uncover the unexpected outcomes of cutting out sugar, and discover the ultimate way to achieve this.
Honey is a sweet food and medicine that nature has gifted us. Its history of medicinal use can be traced back 8,000 years. Sometimes, the therapeutic effects of honey surpass even those of medications, particularly its outstanding antibacterial and antiviral properties.
Honey’s Antibacterial Properties Are Superior to Antibiotics’
“Bacteria quickly learn how to fight off the effects of the antibiotic drugs, but they don’t have honey resistance,” Nural Cokcetin, a research fellow at the Australian Institute for Microbiology and Infection in the science faculty at the University of Technology Sydney, said in an interview with The Epoch Times.
Honey has been used to treat skin problems and wound infections for thousands of years. For instance, ancient Greeks and Egyptians would apply honey topically to the skin to treat wounds and burns. Honey’s effectiveness in treating wounds and eczema was also documented in Persian traditional medicine. Interestingly, the concept of microorganisms such as bacteria and fungi causing wound infections during those historical periods might not have even been known.
The broad-spectrum antibacterial activity of honey has been increasingly recognized and substantiated. In recent years, the prevalence of wound infections caused by antibiotic-resistant bacteria has become a major global health care challenge. Honey can disrupt biofilms that are formed by these resistant bacteria and sometimes penetrate the biofilm structure to eliminate the embedded bacteria. An article published in the journal of the American Society for Microbiology in 2020 reported no documented cases of bacteria developing resistance to honey.
In clinical settings, medical-grade honey has been utilized in various ways. It’s formulated into topical ointments, dressings, and bandages for the treatment of postoperative wound infections, burns, necrotizing fasciitis, nonhealing wounds, boils, venous ulcers, diabetic foot ulcers, eczema, psoriasis, and dandruff. Additionally, it has proven effective in both preventing and treating infections caused by multidrug-resistant microorganisms.
In vitro studies have shown that honey can kill Helicobacter pylori. Furthermore, oral consumption of honey can reduce the duration of bacterial diarrhea.
The antibacterial properties of honey originate from its inherent characteristics and a diverse array of components derived from both nectar-producing plants and bees themselves. (Africa Studio/Shutterstock)
Key Antimicrobial Components
The antibacterial properties of honey originate from its inherent characteristics and a diverse array of components derived from both nectar-producing plants and bees themselves. Additionally, some of these components are formed during the maturation process of honey.
High Viscosity and pH
Honey has a high sugar content, reaching up to 82.5 percent. As a result, it has a viscous and syrupy consistency, exhibiting hygroscopic properties (ability to absorb moisture) and high osmolarity. Bacteria that come into contact with honey undergo dehydration and ultimately perish.
Honey is acidic due to its various organic acids, with a pH value ranging from 3.2 to 4.5. Bacteria can’t survive in honey; their optimal pH typically ranges between 6.5 and 7.5.
Primary Antibacterial Agent: Hydrogen Peroxide
An important enzyme in honey derived from bees facilitates the oxidation of glucose, resulting in the production of hydrogen peroxide.
Hydrogen peroxide is an effective bactericidal agent that can inhibit and kill microorganisms.
Interestingly, water is necessary to induce honey’s hydrogen peroxide production. The maximum level of hydrogen peroxide can be obtained by diluting honey to a concentration of 30 to 50 percent, according to a paper in the International Journal of Microbiology.
Bee defensin-1 is another natural antibacterial component in honey originating from the honeybee’s hypopharyngeal gland.
Bee defensin-1 is an antimicrobial peptide that can kill various bacteria and microorganisms. Specifically, the defensin protein has been shown to create pores within the bacterial cell membrane, leading to cell death.
Additionally, bee defensin-1 can aid in wound healing by stimulating keratinocytes, the most common type of skin cell.
Unique Antibacterial Component in Manuka Honey: Methylglyoxal
Manuka honey is often mentioned in studies regarding the antibacterial effects of various kinds of honey. As a globally recognized honey with high antibacterial potency, it possesses a unique natural component called methylglyoxal (MGO).
After honeybees gather nectar from the flowers of manuka trees, a natural substance that’s found in the trees undergoes spontaneous dehydration, leading to the formation of MGO in the honey.
“The more of [methylglyoxal] there is, the more antibacterial that honey is,” Dee Carter, a professor from the School of Life and Environmental Sciences at the University of Sydney, told The Epoch Times.
MGO exhibits remarkable antibacterial properties by itself. It can alter the structure of bacterial fimbriae and flagella (appendages), and induce damage to their cell membranes, ultimately resulting in cell death.
However, this substance is sensitive to heat. Heating manuka honey to 98.6 degrees F (37 degrees C) can increase the levels of MGO, but heating it to 122 degrees F (50 degrees C) results in the loss of MGO.
Manuka honey possesses a unique compound derived from the manuka tree called methylglyoxal (MGO), which is an antibacterial compound not found in other types of honey.
Honey’s Antiviral Properties May Reduce COVID-19 Complications
Honey not only possesses antibacterial properties but also exhibits antiviral effects, which help protect the body and boost the immune system. Many individuals find relief from symptoms of cold and respiratory illnesses by consuming honey water, and there’s a valid basis for this remedy.
Combats Influenza and COVID-19
MGO can inhibit the growth of enveloped viruses, including the virus that causes COVID-19.
According to a study published in the Eurasian Journal of Medicine and Oncology in 2020, computer simulations revealed that multiple compounds found in honey can bind to the protease of SARS‑CoV‑2, effectively inhibiting its replication.
MGO in manuka honey can also inhibit the replication of influenza viruses, including drug-resistant strains.
Currently, scientists are researching the potential of MGO for managing and treating COVID-19.
Boosts the Immune System
Honey can activate several immune cells, enabling them to eliminate viruses effectively. For example, oligosaccharides that are present in honey can enhance the activity of natural killer cells. Additionally, certain immune cells can “remember” specific viruses during the antiviral response, offering future protection against the same viral invasions.
Honey can promote the proliferative activity of immune cells, thus increasing their quantity.
Promotes Autophagy
Honey can promote autophagy, which is crucial in combating deadly viruses such as SARS‑CoV‑2, which causes COVID-19. It’s an internal cellular process that involves the degradation and metabolism of cells, allowing for the removal and recycling of unwanted or damaged cells and impurities. This process contributes to the generation of healthier cells and promotes recovery.
Honey’s Anti-Inflammatory and Gut-Health Benefits
Inflammation is a common occurrence in bacterial or viral infections. Left unchecked, it can significantly harm the body, potentially leading to life-threatening complications.
Moreover, honey has been suggested to have a dual role in inflammation control: It can downregulate anti-inflammatory factors, thereby reducing inflammation damage, and stimulating the production of inflammatory mediators, promoting wound healing.
In one of Ms. Cokcetin’s studies, honey could reduce inflammation levels in the intestines. She attributes this effect to the abundant presence of phenolic compounds found in honey.
She explained that honey also contains oligosaccharides and polysaccharides, which are complex sugars. These compounds serve as prebiotics, supporting the growth of beneficial bacteria in the gut and inhibiting the growth of harmful bacteria. Additionally, consuming honey promotes the production of short-chain fatty acids in the intestines.
Choosing Honey With Potent Properties
Generally, raw and darker honey varieties tend to have more robust efficacy.
Commercially available honey found on supermarket shelves differs from raw honey sold by beekeepers or at bazaars, as it’s generally pasteurized. Research has shown that heat processing can lower the antioxidant capacity of honey by 33.4 percent. Alternatively, you can also choose honey processed using high-pressure techniques.
Honey comes in a diverse range of colors, from light yellow to amber, dark red, and even nearly black. These colors reflect the varying composition of honey, such as polyphenols, minerals, and pollen. Numerous studies suggest that dark-colored honey generally contains higher levels of phenolic compounds and exhibits more potent activity.
To obtain manuka honey with exceptional antibacterial properties, it’s recommended to opt for pure honey sourced from New Zealand and Australia.
The manuka tree is exclusive to specific New Zealand and Australian regions, and its annual yield is limited. Certain products labeled as manuka honey may contain blends of other honey types, potentially affecting the honey’s properties. Furthermore, some of these products may even include low-quality ingredients such as sucrose.
Opting for single-origin honey sourced exclusively from the manuka tree and checking for its Unique Manuka Factor (UMF) rating is recommended. A higher UMF rating indicates higher levels of substances related to MGO. Some honey products also specify the amount of MGO in milligrams per kilogram.
However, manuka honey with a higher UMF rating may have a strong flavor that some people find pungent.
Ms. Cokcetin recalled that when she began her research on the antibacterial properties of honey, a professor she worked with said, “We’ve already got this great gift from nature that is a solution to superbugs; we just have to figure out how it works and why it works the way it does.”
Researchers are studying how extreme weather and rising temperatures can encourage the spread of drug-resistant infections.
Flooding in Bangladesh in 2020 compromised the safety of water supplies. Credit: Zabed Hasnain Chowdhury/SOPA Images/LightRocket via Getty
In 2021, microbiologist Adwoa Padiki Nartey caught a bacterial infection and her tonsils started to swell. She’d had the same symptoms the year before; both illnesses occurred during one of two annual rainy seasons in Ghana’s capital, Accra, where she lives. In these seasons, she knew, high humidity and moisture spur microbial growth. Doctors prescribed antibiotics but, unlike before, the drugs didn’t work. The bacteria had become resistant.
Over two weeks, her tonsils became increasingly inflamed. “It was painful to drink. Painful to eat. I could barely speak,” says Padiki Nartey, who researches antibiotic resistance at the University of Ghana in Accra. “I was worried.”
To her relief, a final combination of two types of antibiotic worked. Otherwise, the infection could have killed her, she says. “That’s pretty scary.”The fight against antimicrobial resistance
The episode demonstrates how two major threats to human health — climate change and the spread of antibiotic resistance — can intersect. Climate change has led to more frequent heavy rainfall in Ghana, Padiki Nartey says. Those conditions, in turn, encourage bacterial growth, including that of antibiotic-resistant microbes. Flooding also spreads antibiotic-resistant bacteria from sewage systems into people’s homes and drinking water, she adds.
Although researchers have some understanding of how climate change will exacerbate the rise in antibiotic resistance, scientists are now digging into the mechanisms and exploring how large the effect could be, says Pamela Yeh, an evolutionary biologist at the University of California, Los Angeles. “Climate change and antibiotic resistance are two of the biggest health issues of our time,” she says. “Researchers are starting to look at how they link up.”
Climbing resistance
Drug-dodging bacteria are on the rise. In 2020, human blood infections caused by antibiotic-resistant Neisseria gonorrhoea, Escherichia coli and Salmonella bacteria became at least 15% more common than in 2017, according to a report published by the World Health Organization (WHO) in 2022. We are hurtling towards a world where scraping your knee, giving birth or having a simple surgical procedure could kill you, says Yeh. “It’s incredibly frightening,” she says.
The key problem is that antibiotics are often overused or misused to fight infections in people, other animals and plants. Bacteria can develop resistance to drugs through DNA mutations that alter the bacterial cell wall such that antibiotics can’t work, or that confer abilities to break down antibiotics or pump them out of cells. Strains that become resistant can also share antibiotic-resistance genes with other bacteria. If the wrong antibiotics are given to treat infections, or if the right drugs are taken at insufficient doses to kill the microorganisms, then the microbes have more time to multiply and to evolve or spread resistance.
But as in Ghana, changing weather conditions that help bacteria thrive also play a part. And disasters stemming from extreme weather events such as floods, droughts, hurricanes and wildfires can exacerbate the issue because they often reduce access to clean water, leading to unsanitary conditions. When injuries and infections start to rise, more people use antibiotics, in turn pushing up the likelihood of resistance developing.
In a study1 published last October, microbiologist Rita Colwell at the University of Maryland in College Park and her colleagues showed that a deadly hurricane had led to a spike in harmful species of antibiotic-resistant Vibrio bacteria — including a flesh-eating species — in waters off the coast of Florida. The high winds whipped up nutrients from marine sediments that the bacteria thrive on, says Colwell.
Temperature changes
Beyond the physical damage caused by extreme weather, researchers are exploring how rising temperatures that result from climate change might influence antibiotic resistance. In November 2022, microbiologist Lianping Yang at Sun Yat-sen University in Guangzhou, China, and his colleagues reported on the prevalence of three bacteria behind antibiotic-resistant hospital infections that are often severe and can be deadly: Acinetobacter baumannii, Klebsiella pneumoniae and Pseudomonas aeruginosa2.
The researchers looked for links between temperature and antibiotic resistance by comparing bacterial data — collected from people treated in hospitals in 28 provinces and regions across China — with information on average air temperatures in cities in the same regions. The team corrected for factors that could affect rates of drug resistance, including the level of antibiotic consumption, average humidity, yearly rainfall and population density.
They found that, for every 1 °C increase in average air temperature, there was a 14% increase in the proportion of samples containing K. pneumoniae that were resistant to a type of antibiotic called carbapenems. These drugs are usually reserved for treating bacteria that are resistant to all other antibiotics.
Yang and his colleagues also linked a 1 °C increase in average air temperature to a 6% increase in the proportion of samples containing carbapenem-resistant P. aeruginosa. But temperature did not significantly affect the prevalence of drug-resistant A. baumannii.
Widely used antibiotics such as penicillin (in white spots) can have very different effects on related bacterial strains (opaque growths).Credit: John Durham/SPL
These findings support studies from 2018 and 2020 by microbiologist Derek MacFadden at the University of Ottawa and his colleagues3,4. The team found that increased average minimum temperatures were linked to higher rates of antibiotic resistance across 41 states in the United States3 and 28 countries in Europe4. However, the studies do not show a causal link between temperature and antibiotic resistance, says Yang.
One way that gradual increases in average air temperature could be driving up antibiotic resistance is by increasing the growth rate of bacteria, speeding up their evolution, says MacFadden. There is also evidence that bacteria can share genes — including elements that confer antibiotic resistance — more easily in warm conditions than in cold ones5.
Extreme conditions
Another idea is that very high temperatures — which are experienced more often as the globe warms, even though average temperatures rise only slightly — might induce genetic changes in bacteria that help them to resist antibiotics, says Yang.
In a 2018 study, Yeh and her colleagues exposed E. coli, which grew best at 41 °C, either to a temperature of 44 °C or to a range of 12 antibiotics (deliberately given at low doses to inhibit but not kill all the bacteria)6. The researchers tracked how the bacteria responded to these stressors, and found that patterns of gene expression changed in similar ways for both temperature and antibiotic type. In both cases, bacteria responded to the stress by producing more ‘heat-shock’ proteins. These help other proteins to fold correctly and apparently also help bacteria to survive antibiotic attack, says Yeh. “We sometimes call antibiotics that mimic [the effects of] hot conditions ‘hot’ drugs,” she says.
When E. coli was exposed to temperatures as low as 22 °C, the microbes ramped up ‘cold-shock’ proteins — again helping them to produce the proteins needed for survival. Some ‘cold’ antibiotics, such as tetracycline, altered bacterial gene activity in a similar way.COP28 climate summit signals the end of fossil fuels — but is it enough?
The findings suggest that bacteria that have evolved in extremely hot or cold conditions might be more resistant to certain antibiotics, Yeh says. But in a study that is yet to be published, Yeh’s team found that exposure to high temperatures sometimes makes bacteria less resistant to ‘hot’ antibiotics, highlighting that the links between temperature and resistance are likely to be complex, she says.
Extreme temperatures could also contribute to antibiotic resistance by changing how people interact with each other, says MacFadden.
In South Korea, “when we reach the highest temperatures, people tend to stay inside”, says microbiologist Soojin Jang at the Pasteur Institute Korea in Seongnam. Bacteria tend to spread particularly easily between people who are in close contact with each other indoors, she says. That could help to spread resistant strains, and — because more bacteria are multiplying in more people — increase the chance that resistance will emerge.
Over the past year, Jang and her team have been tracking the presence of antibiotic-resistance genes in bacterial samples collected from public toilets, trains and buses. The researchers hope to continue this over the next decade and compare rates of antibiotic resistance with factors such as local air temperature and social behaviour. This could help to reveal how climate change is affecting the spread of resistance, says Jang.
Tackling resistance
Surveillance such as Jang’s could be allied with climate and weather data to help track or predict the spread of antibiotic-resistant bacteria, says Colwell. In a study published last February, she and her colleagues used factors including temperature and rainfall data from satellites to predict when and where cholera outbreaks — caused by the waterborne bacteria Vibrio cholerae — would occur7. Their machine-learning model predicted the risk of cholera outbreaks in various regions across Yemen with an average accuracy of 72%, and up to four weeks in advance.
Combining weather data with surveillance for antibiotic-resistance genes by monitoring waste water could improve forecasts for areas that are at a high risk of antibiotic resistance, Colwell says. It’s an approach her team is working on.
Hurricane Ian caused flooding in the streets of Fort Myers, Florida, in 2022.Credit: Pedro Portal/Miami Herald/Tribune News Service via Getty
Besides improving surveillance, researchers already know what’s needed to reduce the spread of antibiotic-resistant bacteria: improving access to clean water and sanitary facilities, and raising awareness of how to use antibiotics appropriately.
Some efforts have been successful. In Lebanon, for instance, clinician and infectious-disease researcher Souha Kanj says a programme that she and her colleagues started at the American University of Beirut in 2018 educated physicians on how to reduce the use of carbapanem antibiotics in their hospital. In 2020, the proportion of A. baumannii infections that were carbapenem-resistant had fallen to 63%, from 81% at the start of the programme8.
Padiki Nartey, meanwhile, is among those trying to develop new ways to kill drug-resistant bacteria. One particularly promising approach is to use viruses called phages, which infect bacteria, but not human cells.
Global action
Researchers seeking to stem the rise of antibiotic resistance could also learn lessons from climate-change politics, says Steven Hoffman, an international lawyer and health-policy researcher at York University in Toronto, who is also at the Public Health Agency of Canada. Such global threats don’t respect national borders, says microbiologist Saffiatou Darboe at the University of the Gambia in Serrekunda. So, drawing parallels with climate change, countries should agree a global treaty to tackle the issue, Hoffman says.
But stronger action is needed, says Hoffman. In a 2022 study9, he and his colleagues called for a treaty to tackle drug-resistant pathogens — equivalent to the 2015 Paris climate agreement. He wants countries to articulate a single unifying goal, such as a 35% reduction in drug-resistant infections by 2035, and hopes that they will make progress on this when they convene at a UN meeting on antimicrobial resistance in September. The meeting is only the second time that the issue has been the subject of a UN General Assembly.
Growth of bacterial cultures in Petri dishes [Sylvia Suter, University Hospital Basel]
Clinical bacteriology relies on the ability to identify cultured isolates. However, unknown isolates are common in hospital settings. Now, researchers at the University of Basel, have established a new study algorithm—NOVA (Novel Organism Verification and Analysis)—to analyze bacterial isolates that cannot otherwise be characterized using conventional identification procedures.
The team, which has been collecting and analyzing patient samples containing unknown bacterial isolates since 2014, have discovered more than thirty new species of bacteria, some of which are associated with clinically relevant infections.
In this work, the researchers led by Daniel Goldenberger, PhD, at the University of Basel, analyzed 61 unknown bacterial pathogens found in blood or tissue samples from patients with a wide range of medical conditions. Conventional laboratory methods, such as mass spectroscopy or sequencing a small part of the bacterial genome, had failed to produce results for all these isolates. Researchers sequenced the complete genome of the bacteria.
Out of 61 analyzed bacteria, 35 represent potentially novel species. And 27 of 35 strains were isolated from deep tissue specimens or blood cultures. The authors noted that, “Corynebacterium sp. (n = 6) and Schaalia sp. (n = 5) were the predominant genera. Two strains each were identified within the genera Anaerococcus, Clostridium, Desulfovibrio, and Peptoniphilus, and one new species was detected within Citrobacter, Dermabacter, Helcococcus, Lancefieldella, Neisseria, Ochrobactrum (Brucella), Paenibacillus, Pantoea, Porphyromonas, Pseudoclavibacter, Pseudomonas, Psychrobacter, Pusillimonas, Rothia, Sneathia, and Tessaracoccus.”
Most of the newly identified species belong to the Corynebacterium and Schaalia genera, both gram-positive bacilli. “Many species in these two genera are found in the natural human skin microbiome and the mucosa. This is why they are frequently underestimated, and research into them is sparse,” explained Goldenberger. They can, however, cause infections when they enter into the bloodstream—due to a tumor, for example.
The researchers classified the remaining 26 strains as hard to identify. An evaluation of the patient data showed that seven out of the 35 new strains were clinically relevant, meaning that they can cause bacterial infections in humans. “Such direct links between newly identified species of bacteria and their clinical relevance have rarely been published in the past,” said Goldenberger.
One of the hard-to-identify pathogens might be clinically relevant, too. It was found in the inflamed thumb of a patient after a dog bite. A Canadian research group also recently isolated this bacterium from wounds caused by dog or cat bites. “This has led us to assume that it is an emerging pathogen which we need to monitor,” said Goldenberger. The Canadian researchers appropriately named the bacterium Vandammella animalimorsus (animal bite Vandammella) in 2022.
Naming their new species is the next item on the Basel team’s to-do list, too. They are working closely with Peter Vandamme, a professor from the University of Ghent and a specialist in bacteria classification. Two of the bacteria have been named already. One is Pseudoclavibacter triregionum—referring to Basel’s location near the borders of Switzerland, France, and Germany.
Researchers report that a new type of antibiotic has proved its mettle against a deadly superbug.
Acinetobacter baumannii, a bacteria that goes by the nickname CRAB when it becomes antibiotic-resistant, can trigger serious infections in the lungs, urinary tract and blood, according to the U.S. Centers for Disease Control and Prevention. Unfortunately, it’s resistant to a class of powerful broad-spectrum antibiotics called carbapenems.
Now, in a study published Jan. 3 in the journal Nature, researchers from Harvard University and the pharmaceutical company Hoffmann-La Roche discovered that a new type of antibiotic, zosurabalpin, can kill A. baumannii.
Zosurabalpin employs a unique method of action, researcher Dr. Kenneth Bradley, global head of infectious disease discovery with Roche Pharma Research and Early Development, told CNN.
“This is a novel approach, both in terms of the compound itself but as well as the mechanism by which it kills bacteria,” he explained. A. baumannii is a Gram-negative bacteria, meaning it is protected by both inner and outer membranes, making it difficult to attack.
In this study, the scientists first tried to identify and then fine-tune a molecule that could cross those double membranes and eliminate the bacteria.
After years of improving the potency and safety of a number of compounds, the researchers chose one modified molecule.
How does it work? Zosurabalpin prevents the movement of large molecules called lipopolysaccharides to the outer membrane of the bacteria, where they keep the protective membrane intact. This causes the molecules to accumulate inside the bacteria’s cell to the point where the cell becomes so toxic that it dies.
In the study, zosurabalpin worked against more than 100 CRAB samples. It also reduced the levels of bacteria in mice with CRAB-induced pneumonia and prevented the death of mice with sepsis triggered by the bacteria.
Zosurabalpin is now being tested in phase 1 clinical trials, to assess its safety in humans, the researchers told CNN.
Still, the public health threat of antimicrobial resistance remains a huge problem globally due to a lack of effective treatments, Dr. Michael Lobritz, worldwide leader of infectious diseases at Roche Pharma Research and Early Development, told CNN.
In the United States, there are more than 2.8 million antimicrobial-resistant infections each year. More than 35,000 people die as a result, according to the CDC’s 2019 Antibiotic Resistance Threats Report.
Even though zosurabalpin is years away from human use, it’s an extremely promising development, Dr. César de la Fuente, presidential assistant professor at the University of Pennsylvania, told CNN.
“I think from an academic perspective, it is exciting to see a new type of molecule that kills bacteria in a different way,” de la Fuente said. “We certainly need new out-of-the-box ways of thinking about antibiotic discovery, and I think this is a good example of that.”
The only drawback to the discovery is that the modified molecule will work only against the specific bacteria it is designed to kill, the researchers noted.
However, de la Fuente said this new method could turn out to be better than many broad-spectrum antibiotics.
“For decades, we’ve been obsessed with creating or discovering broad-spectrum antibiotics that kill everything,” he noted. “Why not try to come up with specific, more targeted antibiotics that only target the pathogen that is causing the infection and not all the other things that might be good for us?”
The gut microbiome, i.e., the community of bacteria and other microorganisms living in the human gut, has been implicated both directly and indirectly (mediating the effects of diet) on human health [1, 2]. The associations between gut microbiome composition and disease status have been widely reported, while recent studies have demonstrated a role for the gut microbiome in influencing remote organs, mucosal, and immune function [3, 4]. Considerable effort is currently focused on understanding the natural history of microbiome development in humans in the context of health outcomes, in parallel with improving our knowledge of microbiome–host molecular interactions. These efforts ultimately aim to develop effective approaches to rehabilitate perturbed human microbial ecosystems as a means to restore health and prevent disease. This review details the role of the gut microbiome in chronic diseases (Fig. 1) and ways it can be modulated for the management or prevention of chronic conditions.
Fig. 1: Schematic representation of the association of the composition of the gut microbiome and gut-derived metabolites with chronic diseases.
Differences in gut microbiome composition and function have been associated with a variety of chronic diseases ranging from gastrointestinal inflammatory and metabolic conditions to neurological, cardiovascular, and respiratory illnesses.
The aim of this narrative review is to describe the associations between gut microbiome composition and various types of chronic diseases and to discuss the links to habitual diet and dietary components (Table 1).Table 1 Summary of key findings outlining the role of the gut microbiome in chronic disease.
The pathogenesis of autoimmune diseases (AIDs) is not only attributed to genetic susceptibilities but also environmental factors, among which, a disturbed gut microbiota has attracted increasing attention. Compositional and functional changes of gut microbiota have been reported in various autoimmune diseases, and increasing evidence suggests that disturbed gut microbiota contributes to their immunopathogenesis.
Rheumatoid arthritis (RA)
Rheumatoid arthritis (RA) is a systemic autoimmune inflammatory condition that manifests in joint damage. It was recently demonstrated that different environmental factors are involved in the development of both intestinal/oral dysbiosis and arthritis onset and outcome, among which the most relevant are diet, smoking, and infections [5, 6]. The observation that germ-free mice are protected from development of experimental arthritis [7] suggests a possible role for the microbiome in the pathogenesis of this disease. The composition of the gut microbiota in RA patients free of therapy is severely altered compared to healthy controls. Chen et al. reported that compared with healthy controls, patients with RA show decreased gut microbial diversity, which correlates with autoantibody levels and disease duration [8]. In addition, at the compositional level, patients with RA show an increased abundance of Prevotella species, including Prevotella copri [8, 9], and recent preclinical phase studies on RA patients in European countries were shown to harbor a high abundance of this species in the intestine, suggesting that dysbiosis precedes the development of arthritis [10]. In contrast, Faecalibacterium, which is generally recognized as a beneficial microbe, is decreased in RA patients. Furthermore, the relative abundance of Collinsella was found to be increased in RA patients [11]. Interestingly, inoculation of Collinsella into collagen-induced arthritis (CIA)-susceptible mice induces severe arthritis. In vitro experiments showed that Collinsella Aerofaciens increases gut permeability and induces IL-17A expression, a key cytokine involved in the pathogenesis of RA, suggesting that Collinsella is a candidate arthritogenic bacterium in the human intestine [11]. In summary, Prevotella copri [12] and Collinsella are the dominant gut microbiota in patients with early RA and may be involved in its pathogenesis.
Recent links have been made between dietary intake of short-chain fatty acids (SCFAs) and autoimmune arthritis in mice, wherein SCFAs play an important role in the suppression of inflammation in RA [13, 14]. Mice deficient for SCFA receptors showed exacerbated inflammation in modes of RA [14]. Butyrate, one of the most abundant SCFAs, acts as an endogenous histone deacetylase (HDAC) inhibitor and has been shown to decrease inflammation in animal models of RA and other inflammatory diseases [15]. A recent study has revealed a role for intestinal barrier function, and specifically for zonulin, a peptide that controls epithelial tight junction permeability, in regulating the onset of joint disease in mice with collagen-induced arthritis (CIA) and potentially also in patients with RA [16]. Increased levels of zonulin have been associated with leaky intestinal barrier, dysbiosis, and inflammation. Restoration of the intestinal barrier in the period before clinical arthritis, either by dietary supplementation with the SCFA butyrate or pharmacological agents such as a zonulin antagonist may help delay disease onset and reduce the severity of RA.
Type 1 diabetes (T1D)
In humans, intestinal microbiota alterations, including loss of bacterial diversity, preceded the onset of metabolic symptoms associated with T1D [17]. Previous studies have linked several facets of gut health with the onset of T1D in humans and animal models [18,19,20]. In animal models, antibiotic-induced dysbiosis, altered microbial lipid metabolism, and suppressed enteric Th17 and T-reg cell populations have led to increased incidence of T1DM-like disease in mice [21]. Various human-based studies have also reported altered intestinal microbiota in connection to T1D from several ethnic groups [1720, 22,23,24]. The common findings from these studies include increased numbers of Bacteroides species, and deficiency of bacteria that produce SCFAs [25, 26] in cases of T1D. Specifically, the butyrate producer Faecalibacterium prausnitzii has been found to be decreased in abundance in children with diabetes-related autoantibodies [26].
In addition, increased intestinal permeability [22] and decreased microbial diversity [17] before T1D diagnosis have been reported. A multicenter case–control study of 783 children, showed that the microbiomes of healthy children contained more genes that were related to the fermentation and the biosynthesis of SCFAs, but these were not consistently associated with the abundance of particular taxa across geographically diverse clinical centers, suggesting that microbial function rather than composition was associated more with T1D [27].
The protective effects of SCFAs in T1D have been elucidated in animal studies that have looked at nonobese diabetic (NOD) mouse models [28]. For example, NOD mice fed specialized diets resulting in high bacterial release of acetate and butyrate were almost completely protected from T1D [28] mainly via the immune-modulating effects of SCFAs. In animal models of autoimmune diabetes, increased gut permeability precedes the development of diabetes, and environmental factors that modulate the permeability thereby modulate the incidence of the disease. Gut permeability appears to be an important factor in the relationship between the intestinal microbiota and the development of T1D. Both gut permeability and inflammation have been linked to the development of T1D in humans [26, 29].
Atopic eczema
Atopic eczema, a chronic inflammatory skin disorder, is most prevalent in early childhood. The pathogenesis of eczema has been attributed to skin-barrier dysfunctions, immune dysregulation as well as environmental–host–microbial interactions [30]. Environmental factors and modern lifestyle trends have been shown to indirectly contribute to pathogenesis of the disease through modulation of the gut microbiome [31].
Previous studies have found that the gut microbiome in early life is associated with age of onset, severity, and remission of atopic eczema [32, 33]. However, the association between gut microbiome diversity and atopic eczema development remains contradictory with few studies reporting an inverse relationship between gut diversity and the severity of atopic eczema [34, 35]. The development of atopic eczema may instead be driven by the interactions between specific gut microbiome signatures, the immune system, and the host. Specifically, in patients with atopic eczema, the proportion of Clostridia, Clostridium difficile, Escherichia coli, and Staphylococcus aureus in the gut microbiome is higher than in healthy controls, whereas that of Bifidobacteria, Bacteroidetes, and Bacteroides is decreased [34, 36,37,38]. The abundance of Clostridia and Escherichia coli in the intestine may be associated with atopic eczema via eosinophilic inflammation [36]. Butyrate-producing bacteria, such as Coprococcus eutactus, are increased in infants with milder atopic eczema or healthy infants than in those with severe atopic eczema [34]. Furthermore, fecal samples from patients with atopic eczema showed decreased levels of butyrate and propionate, which have anti-inflammatory effects. This is likely a consequence of a dysbiosis in the SCFA producer F prausnitzii that reduces the number of high butyrate and propionate strains [38].
It is evident that the compositional and proportional differences in the gut microbiome are associated with the development of AID via an immunomodulatory effect of the gut microbiome. The gut microbiome may contribute to the development, persistence, and severity of AID via immunologic, metabolic, and neuroendocrine pathways. It is however still unclear whether compositional changes in the gut microbiome precede the development of atopic eczema, and thereby shift the immune system and disrupt the gut epithelial barrier, making it amenable to the development of atopic eczema.
Atopic asthma
The rapid increase in asthma prevalence in industrialized nations over the past several decades cannot be explained by genetic risk factors alone and is thought to be related to altered environmental exposures associated with Western lifestyles. Early life is the most important period during which microbiota dysbiosis in the gut may lead to the development of many respiratory diseases, as the gut microbiota has a significant influence on immune-cell maturation and resistance to pathogens [39]. A number of validated epidemiological observations have implicated early-life environmental exposures in increased risk for childhood asthma. Many of these exposures are known to shape the nascent gut microbiome, including cesarean birth [40], antibiotic use [41], formula feeding, and other environmental factors including airborne toxins [42]. Further evidence for an intricate relationship between environmental exposure, the gut microbiome, and allergic airway disease comes from an expanding body of work, particularly those utilizing experimental animal models. For example, treating neonatal mice with antibiotics was shown to diminish gut microbial diversity, alter metabolite profiles, exacerbate immune cell response, and increase susceptibility to allergic lung inflammation [43, 44]. Furthermore, supplementation with SCFAs ameliorated airway inflammation in these mice, with the mechanism attributed to decreased activity of immune-modulating markers such as T cells, IL-4-producing CD4+ T cells, and reduced levels of circulating IgE [44].
Observational studies in humans have identified Proteobacteria to be the most dominant phylum overrepresented in patients with asthma compared with nonasthmatic volunteers across several studies [39]. The Proteobacteria phylum is represented by potentially pathogenic bacteria, including those that belong to the genera Haemophilus, Moraxella, and Neisseria [39].
Asthma pathogenesis and severity are linked with pro-inflammatory mechanisms. The effects of the gut microbiota on asthma are at least partially mediated by bacterial metabolites, which may influence immune responses in distal parts of the body. The most known metabolites with demonstrated protective properties in human airway inflammation are SCFAs. Children with high amounts of butyrate and propionate in feces at 1 year of age have significantly less atopic sensitization and are less likely to have asthma between 3 and 6 years [45]. In addition, soluble fiber may ameliorate the effects by exerting anti-inflammatory action via SCFAs binding to associated G-protein-coupled receptors (GPCRs) [46, 47].
Of recent interest are studies showing that gut bacteria in humans are able to produce other metabolites with pro- and anti-inflammatory potential, such as biogenic amines (including histamine) [48] and oxylipins such as 12,13-diHOME [49]. The number of histamine-secreting bacteria is significantly higher in fecal samples of asthma patients compared with nonasthmatic volunteers [50]. Furthermore, the number of histamine-secreting bacteria correlates with disease severity.
Although the mechanisms mediating communication between the gut and lungs are still unclear, it has been suggested that epithelial cells, other structural cells, and immune cells absorb signals from the gut endothelium to form a local cytokine microenvironment, which leads to changes in immune responses at distal sites [51]. Specifically, SCFAs derived from gut bacteria have inhibitory effects on proinflammatory responses in the lungs.
Gut microbiome–gut inflammation/bowel disorders
Irritable bowel syndrome (IBS)
IBS is generally characterized by abdominal pain, discomfort, and altered bowel habits. Although the etiology is multifactorial, recent understanding of the pathophysiology of IBS has revealed that variations in the normal gut microbiota may have a role to play in the low-grade intestinal inflammation associated with the syndrome [52]. Microbial dysbiosis in the gut is thought to be involved in IBS pathogenesis [53] and a recent study revealed a clear separation between the gut microbiota of patients with IBS and that of the controls. IBS was characterized by an increase in Firmicutes and, more specifically, in the numbers of Ruminococcus, Clostridium, and Dorea, in addition to a marked reduction of beneficial microbes such as Bifidobacterium and Faecalibacterium spp. [54]. Furthermore, systematic reviews have demonstrated the potentially harmful microbiota in patients with IBS, including phylum Proteobacteria, family Enterobacteriaceae (phylum Proteobacteria), family Lactobacillaceae, and genus Bacteroides (phylum Bacteroidetes). The Enterobacteriaceae family contains several pathogenic bacteria; for instance, Escherichia, Shigella, Campylobacter, and Salmonella [55]. These could either suggest previous intestinal infection in such patients or a change in the intestinal environment. By-products from these potentially harmful bacteria have been associated with some of the classical symptoms of IBS, including abdominal pain, bloating, and diarrhea [55].
The most consistent finding of a potentially “protective” bacterial group in IBS patients was found in the group of uncultured Clostridiales. Although the association is not causal and the mechanism of a protective effect in IBS symptoms is unclear. The genus Faecalibacterium, especially Faecalibacterium prausnitzii, which belongs to the similar order as the uncultured Clostridia, has been associated with maintaining gut mucosal health. This bacterium was considered as the main butyrate-producing and anti-inflammatory organism [56] and reduced IBS symptoms via mediation of IL-17 expression in a rat model [57, 58], as well as maintained gut-barrier integrity [59]. Furthermore, the genus Bifidobacterium was decreased significantly in IBS patients regardless of IBS subtype. Therefore, it was another promising potential genus in ameliorating IBS symptoms. A systematic review of probiotics in IBS has highlighted that Bifidobacterium-containing interventions reduce IBS symptoms, which are not seen in products that contain Lactobacillus alone [60].
Management of IBS symptoms can be achieved by the restriction of fermentable oligosaccharides, disaccharides, monosaccharides, and polyols (FODMAP). Clinical studies have now shown the clinical efficacy of the low FODMAP diet in reducing IBS symptoms [61]. One consequence of this dietary intervention is its impact on gut microbiome composition as FODMAPs can modulate microbial composition and microbial metabolite production [62] and not all IBS subjects respond and there may be challenges with implementing a low FODMAP diet. Given the role of the microbiome in metabolizing poorly absorbed carbohydrates that depend on a person’s microbiome composition, that there has been increasing attention on the potential role of the gut microbiome in predicting low FODMAP diet efficacy [62] and recent data suggest that there are potential baseline differences in microbiome activity and composition, which can distinguish between low FODMAP diet responders and nonresponders [63].
Inflammatory bowel disease (IBD)
Long-lasting inflammation and ulceration of the colon are predominantly the main characteristics of IBD, which are features of Crohn’s disease (CD) and ulcerative colitis (UC). IBD, encompassing both ulcerative colitis and Crohn’s disease, is characterized by chronic and relapsing inflammation of the GI tract. The onset of both conditions is, in general, not thought to be due to a single causal organism but by a general microbial dysbiosis in the gut [64]. A role for gut microbes in the manifestation of IBD has been indicated by a number of studies and the gut microbiota are thought to be essential components in the development of mucosal lesions [65]. The alterations in the composition and functionality of the gut microbiota in IBD compared with non-IBD controls have been shown previously. In general, microbial dysbiosis in IBD is characterized by a decrease in diversity and stability of the microbiota [66,67,68,69]. Specifically, a decrease in Firmicutes and an increase in Proteobacteria taxa is the most consistent outcome from IBD microbiome studies. Furthermore, a common signature of microbial dysbiosis among IBD patients, especially in (active) CD, is the decreased abundance of Firmicutes bacteria belonging to the families Ruminococcaceae and Lachnospiraceae as opposed to healthy control samples [70,71,72,73]. Both families are important functional members of the human gut microbiota since most butyrate-producing bacteria from the human gut belong to them. Therefore, depletion of these bacterial families in IBD can be linked to the observed disturbance on a functional level, including a lower butyrate-producing capacity of the IBD microbiota [74]. In addition, a metagenomic and proteomics study in ileal CD microbiota demonstrated an underrepresentation of genes for SCFA production and a decrease in metagenomic reads and proteins of important butyrate producers Faecalibacterium prausnitzii and Roseburia sp. [75].
Butyrate has therapeutic potential in IBD because it serves as the main energy source for colonocytes, enhances epithelial barrier integrity, and inhibits inflammation. Recent observational and interventional studies have explored an alternative approach of probiotics, which involves the consumption of butyrate-producing bacteria to increase in situ butyrate production [76,77,78]. This may suggest that targeting microbial dysbiosis by supplementing butyrate-producing bacteria could restore gut homeostasis and health in IBD.
Gut microbiome composition and cardiometabolic diseases
Cardiovascular disease
The alterations in gut microbiota composition and the metabolic potential of gut microbiota have been identified as a contributing factor in Cardiovascular Disease (CVD) development [79]. Recently, mechanistic links between the gut microbiota and the severity of myocardial infarction have been reported in rats [80, 81]. Hypertension is the most prevalent modifiable risk factor for CVD. However, results from human studies remain scarce and conflicting. In the TwinsUK cohort [82], self‐reported hypertension was not related to 68 various microbiota markers. Furthermore, in the CARDIA study of 529 individuals an increase in alpha diversity resulted in a modest decrease in objectively measured systolic blood pressure (BP) [83]. More recently, the association between the gut microbiome and hypertension was studied on the well-phenotyped population of 6953 individuals as part of the FINRISK study [84] using standardized BP measurements. Although the associations between overall gut taxonomic composition and BP were weak, individuals with hypertension demonstrated changes in several microbiota genera, with most of these genera belonging to the Firmicutes phylum. Interestingly, most of the negative associations with BP belonged to the Lactobacillus species, specifically the greater abundance of Lactobacillus paracasei, a known probiotic that was associated with lower mean arterial pressure and lower dietary sodium intake which has also been reported previously by Wilck et al. [85].
Metabolism of certain dietary components such as choline and carnitine, which comprise a large component of a Western diet, has been shown to increase risk of cardiovascular disease mainly via the metabolism of these compounds to produce trimethylamine oxide (TMAO) [86, 87]. Gut-derived metabolites such as SCFAs have been recently shown to modulate blood pressure [88]. The suggested mechanism of SCFAs to modulate blood pressure is via the host G-protein coupled receptor (GPR) pathways that impact renin secretion and blood pressure regulation [88]. A series of studies using GPR41 knockout mice further supports involvement of these receptors in blood pressure control. This suggests that the gut microbiota are potentially intertwined functionally to control blood pressure, and their dysfunctions could be associated with hypertension. A recent study found that a Mediterranean diet which typically encompasses a high fiber diet followed by fasting was shown to reduce systolic blood pressure, need for antihypertensive medications, body-mass index at three months post intervention compared to following a Mediterranean diet alone [89]. These data highlight fasting followed by a shift to a health-promoting diet as a promising non-pharmacological intervention for patients with hypertension and other cardiometabolic diseases.
In rodent model studies, the administration of probiotics such as Lactobacillus plantarum and Lactobacillus rhamnosus was associated with improved ventricular function and attenuated heart failure after myocardial infarction [80, 90]. Furthermore, a meta-analysis demonstrated a significant decrease in blood pressure in patients treated with probiotics [91]. Lastly, supplementation of atherosclerosis-prone mice with A. muciniphila, a mucus colonizer protected against atherosclerosis development induced by feeding a Western diet [92]. These observations may suggest that probiotics use, in combination with standard medication, could offer additional benefits in the prevention and management of cardiovascular events.
Type 2 diabetes
Adults with T2DM have an altered gut microbiota composition compared to healthy controls. However, it is unclear whether these changes represent a cause or an effect and required further investigation. Among the commonly reported findings, the genera of Bifidobacterium, Bacteroides, Faecalibacterium, Akkermansia, and Roseburia were negatively associated with T2D, while the genera of Ruminococcus, Fusobacterium, and Blautia were positively associated with T2D [93, 94]. A large metagenome-wide association study found a moderate degree of gut dysbiosis in patients with T2DM [95]. Of the identifiable bacterial species, the control samples were enriched in various butyrate-producing bacteria and increased abundance of Lactobacillus spp., while patients with T2DM were characterized by an increase in certain opportunistic pathogens, such as a number of Clostridium spp. [95]. Overall, T2DM was associated with a reduced abundance of butyrate-producing bacteria and SCFAs, in particular butyrate which has been linked with insulin sensitivity [9, 96, 97]. The link between SCFAs and insulin sensitivity lies in the ability of SCFAs to trigger the secretion of GLP-1 by intestinal L-cells via specific G protein receptors (GPR41, GPR43) which has a substantial impact on pancreatic function and insulin release, as well as central effects regulating appetite [98, 99].
In addition to SCFAs, many other metabolites have been implicated in the role of gut microbiome composition and risk to T2DM. We have in fact reported reproducible associations between serum metabolites of microbial origin and gut microbiome alpha diversity in 1018 individuals from the Twins UK cohort [100]. The circulating levels of these metabolites were combined into a microbiome-metabolite score using a linear combination. Importantly the association of the microbiome metabolite score with cardiometabolic traits was stronger than the association between gut microbiome diversity and the same clinical traits. Specifically, the microbiome metabolite score was associated with prevalence and incidence of T2DM in the discovery cohort and this result was replicated in a multi-ethnic independent US cohort (n = 1522) showing the relevance of microbiome-derived metabolites in predicting risk and supporting their role as the mechanistic link between microbiome composition and health [100].
Non-alcoholic fatty liver disease (NAFLD)
Non-alcoholic fatty liver disease (NAFLD) and the more advanced stage non-alcoholic steatohepatitis (NASH) are common comorbidities of obesity and T2DM with an increasing burden for society [101]. There is an increasing body of evidence linking the gut-liver axis to the development of NAFLD. Gut dysbiosis is directly related with increased intestinal permeability as a consequence of epithelial barrier deterioration, tight junctions’ alteration and bacterial translocation causing endotoxemia, which might reach and damage the liver through the portal vein [102,103,104]. Several studies have revealed alterations in the gut microbiome in people with NAFLD compared with healthy controls. We and others have reported a decrease in both alpha and beta diversities in NAFLD and its more advanced stages compared to healthy controls [105, 106]. Case control studies have reported consistent altered microbial signatures at phylum level such as increased Proteobacteria [107,108,109,110], at family level such as increased Enterobacteriaceae and genera such as an increase in the abundance of Collinsella sp, Escherichia, Dorea and a decrease in the abundance of Coprococcus, Eubacterium, Faecalibacterium and Prevotella [107, 111]. Similar microbial signatures were found in NASH as outlined in this recent review [112]. Although these initial results suggest a measurable contrast in microbial signatures between individuals with hepatic steatosis and controls, there are, however, large discrepancies found across studies with varying results in the literature. However, a common microbial signature that has been associated with other metabolic diseases is the reduction of levels of the butyrate producer F. prausnitzii in the more advanced forms of NAFLD [106, 113].
Several hypotheses have provided mechanistic insights into the pathways of how the gut microbiota might contribute to NAFLD development and its disease progression. Specific microbial species such as Collinsella sp. have been shown to metabolize bile acids to oxo-bile acid intermediates which may increase intestinal permeability and contribute to the development of NAFLD [114, 115]. Increased intestinal permeability may lead to lipopolysaccharide (LPS) release to the host, which can trigger tissue and systemic inflammation. Furthermore, the action of microbially-produced metabolites (including TMAO, choline or ethanol) and bile acid signaling can also affect host immunity [108, 116, 117]. Interestingly, unlike other diseases, SCFAs appear to prevent but also promote the development of NAFLD and NASH depending on the signaling pathway or mechanism they activate. Amongst the SCFAs, acetate was found to be enriched in fecal samples from adults with more advanced stages of NAFLD (i.e. fibrosis) [110], whereas butyrate and propionate were enriched in fecal samples from adults with mild or moderate NAFLD. This may suggest that levels and subtypes of SCFAs correlate with the severity of fatty liver disease. Since each SCFA exerts specific and somehow different metabolic effects, assessing their balance both at the fecal and systemic level in patients and after a dietary intervention using different substrates may help provide more information on their overall role in NAFLD development, exacerbation or improvement.
On the basis of the above, advances in discovering gut microbiota or microbiota-related metabolite signatures could be used as noninvasive diagnostic tools in NAFLD and its etiology.
Chronic kidney disease (CKD)
There has been a growing interest in studying the composition of the gut microbiota in patients with CKD as well as the mechanisms by which gut dysbiosis contributes to CKD progression. The influence of the gut microbiota on the gut–kidney axis acts in a reciprocal way with either CKD significantly modifying the composition and functions of the gut microbiota. Alternatively, the gut microbiota is able to manipulate the processes leading to CKD onset and progression through inflammatory, endocrine, and neurologic pathways. Therefore, understanding the complex interaction between these two organs may provide novel interventions to prevent the progression of CKD by targeting the gut microbiota.
The existence of intestinal microbiota alterations such as decrease of microbial richness, diversity, and uniformity has been related to CKD [118]. Patients with CDK show a lower colonization of Bifidobacterium sp, Lactobacillaceae, Bacteroidaceae, Akkermansia, and Prevotellaceae genera, and higher intestinal levels of Enterobacteriaceae, particularly Enterobacter, Klebsiella, and Escherichia, and also increased levels of Enterococci and Clostridium perfringes [119, 120]. The decrease in the abundance of Akkermansia muciniphilla, an important probiotic, in patients with CKD negatively correlated with plasma IL-10 levels, suggesting that an altered microbiota in CKD may promote chronic systemic inflammation [121]. This chronic systemic inflammation state represents a major risk factor for CKD progression.
CDK patients are characterized by decreased consumption of dietary fibers that are required for SCFA production. Lack of dietary fibers leads to increased amino nitrogen, which can be transformed into uremic toxins by the gut microbiota [122]. Patients with CDK are characterized by an imbalance between fermentative and proteolytic microbiota in favor of the latter. The imbalance in favor of proteolytic species is related to detrimental effects and also has a fundamental role in the progression of CKD [123]. Moreover, the main SCFAs especially butyrate, were found to be reduced in the feces and serum of patients during CKD development [123]. Further research is however needed to determine whether increasing levels of circulating SCFAs would provide any direct clinical benefit in patients with CDK.
Several experimental and clinical studies have shown the beneficial effects of prebiotic, probiotic, and synbiotic supplementation on gut microbiota–renal axis [124]. These have emerged as a potential therapeutic intervention to restore the imbalance of the gut microbiota, reduce inflammation or oxidative stress markers and modulate gut-derived uremic toxins, such as P‐cresyl sulfate (PCS), indoxyl sulfate (IS), and trimethylamine N‐oxide (TMAO), which have been implicated in the progression of CKD [125,126,127].
Mental health disorders
Gut microbes have been shown to also influence neurological functions and these links have been termed the “gut−brain axis” [128]. The gut microbiota communicates with the brain through three major pathways—the neural pathway (vagus nerve, enteric nervous system), the immune pathway (cytokines), and the endocrine pathway (HPA axis, gut hormones). Impaired functioning of this connection can lead to manifestation of mental disorders. Common gut microbial species belonging to the phylum Firmicutes and Actinobacteria, and the genera Bacteroides and Bifidobacterium, may be contributing to mental health disorders [129]. Different mental disorders are characterized by the differential gut microbial community that is generally predominated by a bacterial genus or a family as summarized in Table 2. Notably, in some of the disorders, there is an over-abundance of specific microbial species. For example, abundance of Lactobacillus sp. and Bifidobacterium sp. in disorders like schizophrenia (SCZ) [130] and attention-deficit hyperactivity disorder (ADHD) [131], respectively. The probable reason for this can be attributed to the complexity of the gut microbiota.Table 2 Studies (or reviews) on non-dietary interventions targeting the gut microbiome.
Gut microbiota modulates the gut−brain axis via numerous direct and indirect ways. This includes maintaining gut permeability by modulating the integrity of tight junctions in the gut epithelium, producing a wide variety of metabolites including neurotransmitters, SCFAs, and amino acids. These gut-derived metabolites can affect the central nervous system by acting locally on the enteric nervous system or enter circulation and affect the brain. Furthermore, alteration in the levels of gut microbial metabolites, such as SCFAs, ammonia, tryptophan, and histamine have shown to be associated, directly or by breaking down into neuroactive catabolites with various neurological conditions like Parkinson’s disease (PD) [132], anorexia nervosa (AN) [133], Alzheimer’s disease (AD) [134], autism spectrum disorder (ASD) [135], and chronic stress and depression [136]. However, whether this disruption of homeostasis in the mental health disorders is the cause or effect of the alterations in gut microbiota and its functions is not emphatically clear and requires more studies.
Further, considering the evidence, several studies have been undertaken to specifically target the gut microbiota through different therapeutic interventions including administration of pre- and probiotics (psychobiotics) to treat mental health disorders and/or their symptoms [137, 138]. Human-intervention studies with psychotics such as the probiotic combinations of lactobacilli and Bifidobacteria showed significant reduction in psychological distress [139], improved cognition and communication among patients with AD [140] and ASD [141], and improving symptoms among PD patients [142]. Based on the promising effects of psychobiotics on modulating the gut−brain axis, more clinical trials are currently being conducted to identify the bacterial strains as potential candidates for treatment of mental disorders.
The gut microbiome as a therapeutic target in chronic diseases
Given the tight interplay between enteric microbial symbionts and host immunity (Fig. 1), efforts have focused on implementing various strategies targeting the gut microbiota to manage or prevent chronic diseases. Clinical approaches to modify gut microbiota generally focus on depleting overabundant members or the overall microbial load using antibiotics or antifungal agents, modulation through diet, or supplementation with live microbes (single or mixed species). There are a variety of proposed therapeutic approaches, such as dietary modifications, prebiotics, probiotics, and TMAO-synthesis inhibitors, to target the gut microbiome. More recently, fecal microbial transplantation (FMT) has been used in a range of infectious, neurological, and GI conditions, with promising outcomes (Table 3).Table 3 Summary table of key findings.
More recently, the composition of the gut microbiome has been implicated in predicting the severity of COVID-19 possibly via its immune-modulatory properties. More specifically, gut commensals with known immunomodulatory potential, such as Faecalibacterium prausnitzii, Eubacterium rectale, and Bifidobacterium adolescentis, were found to be significantly under-represented in patients with COVID-19 compared with healthy controls and were also associated with disease severity after taking account of antibiotic use and patient age [143]. Furthermore, the microbial imbalance found in the COVID patients was also associated with raised levels of inflammatory cytokines and blood markers of tissue damage, such as C-reactive protein and certain enzymes. Based on these findings, increasing the levels of beneficial gut species depleted in COVID-19 could serve as a novel avenue to mitigate severe disease, highlighting the importance of the gut microbiota in the pathophysiology of COVID-19.
Despite encouraging evidence from animal models in which inflammatory conditions were successfully treated via gut microbiota manipulation, data from human trials is less conclusive. Findings from recent work showed that the ability of an introduced microbe to successfully colonize the gut depends on the structure of the resident gut microbiota [144], a factor that has so far been overlooked in microbial intervention-based clinical trials and that might explain discrepancies in animal models and human trials. Nonetheless, intervention with a multispecies consortium of bacteria has shown to be effective in the maintenance of remission in patients with UC but not in those with CD [145]. Meta-analysis of intervention trials in which various combinations of bacterial strains were administered to adults with T2DM showed moderate improvements in hyperglycemia [146]. Moreover, probiotic supplementation during the first 27 days of life reduced the risk of islet autoimmunity in a large multicenter prospective cohort study of children at high genetic risk for T1DM, compared with no supplementation or supplementation later in infancy [147].
Intervention studies involving live microbial supplementation have also shown encouraging results, although more attention to microbial strain selection based on functional attributes, defined timing or duration of supplementation, and/or tailoring of the supplemented organisms to the endogenous gut microbiome of the recipient may significantly improve the efficacy in future studies. Ongoing studies are focused on understanding the basis of microbe-microbe interactions in order to identify discrete gut microbiomes that more readily respond to specific microbial interventions.
There is also a great need for tailored interventions that consider the microbial individuality of the recipient in order to prevent or manage chronic diseases. Such approaches are likely to result in a transition away from historically used probiotic strains, which are frequently poorly adapted to the enteric microenvironment, toward personalized multispecies microbial consortia sourced from healthy human enteric ecosystems. To further enhance the efficacy of microbial supplementation, nutritional support in the form of targeted dietary modifications, tailored to the specific substrate requirements [148], should also be considered as discussed in the section below. Such an integrated approach to microbiome-based therapeutics, built on independent observations in the field of gut microbiome research, may offer more effective, predictable, and sustainable microbial restitution in cases of chronic disease in which microbiome perturbation and functional gene loss are prominent features.
FMT has proven effective in >90% of patients with recurrent antibiotic-resistant C. difficile infection [149]. The success rate of FMT administration for other chronic intestinal inflammatory conditions such as IBD, however, is more modest, with clinical remission being less predictable [150]. More recently, FMT has been applied to chronic inflammatory conditions affecting organs distal to the GI tract. FMT from lean donors improved insulin sensitivity in men with obesity and metabolic syndrome, and this improvement was linked to changes in plasma metabolites, in particular increased gamma-aminobutyric acid (GABA), tryptophan, and phenylalanine [151].
Effects of a healthy diet are mediated via the gut microbiome
The most obvious way of targeting the gut microbiome is through dietary modifications. Already, several controlled clinical dietary intervention studies targeting the human gut microbiota have been reported [2, 152]. For example, diets rich in fiber were shown to significantly improve glucose control and promote a healthier metabolic profile in T2DM patients as well as reduce the risk of coronary heart disease [153]. Other studies showed marked interindividual variation in postprandial glycemic responses after consumption of identical meals [154, 155]. The consideration of microbial composition alongside known disease-risk factors (i.e., body mass index, fasting glucose) enabled accurate glucose-response prediction, thus allowing the design of more effective, personalized diets for improved glycemic control [155] in these studies.
Some of these same studies have recently investigated the links between gut microbiome structure, habitual diet, and blood cardiometabolic markers [156] in a cohort of 1100 healthy individuals from the United States and the United Kingdom. This study reported that microbial diversity and composition were highly correlated with both fasting and postprandial biomarkers. The strongest associations were with high-density lipoprotein cholesterol (HDLC), which positively correlated with microbial species richness. Cardiometabolic markers, which have in the recent past been shown to be strongly predictive of incident diabetes and heart disease, such as glycoprotein acetyl (GlycA) were also associated (positively or negatively) with microbiome richness. One of the most striking findings of the study was that gut microbiome composition is a better predictor of postprandial triglyceride and insulin concentrations than of glucose levels. The authors also reported that the diversity of healthy plant-based foods in habitual diet shapes gut microbiome composition. Approximately 10% of the variance in gut microbial composition could be explained by variation in healthy eating patterns [156].
Bacterial species were found to segregate into groups associated with more and less healthy plant- and animal- based foods. In addition, the authors reported a microbial signature of cardiometabolic health levels consistent across diet, obesity indicators, and cardiometabolic risks. The authors reported a consistent set of microbial species that strongly linked to all of the following: healthy eating patterns, indicators of obesity and cardiometabolic health, and fasting and postprandial circulating metabolites connected with cardiometabolic risk [156]. Other much smaller studies have also reported that dietary habits are strongly correlated with gut microbiome composition, disease status, and SCFA production [129].
Although a key weakness of the above study is its cross-sectional observational nature, consistent changes in gut microbiome composition and health outcomes have been recently reported by an international consortium investigating the effect of 12 months of Mediterranean diet on 612 elderly individuals from five countries [157]. Shankar Ghosh et al. [157] reported significant differences in the association patterns of bacterial taxa that increased (DietPositive) or decreased (DietNegative) in response to the Mediterranean diet intervention. The DietPositive bacteria had consistent negative associations (significantly lower than the DietNegative bacteria) with the pro-inflammatory circulating markers hsCRP and IL-17 levels as well as with functional measures associated with increased frailty. In contrast, DietPostive bacterial taxa showed positive correlations with measures of improved cognitive function, hand-grip strength, and two of the anti-inflammatory cytokines. DietPositive species included butyrogenic taxa such as F. prausnitzii, Eubacterium and Roseburia, whereas DietNegative bacteria included Ruminococcus torques, Collinsella aerofaciens, Coprococcus comes, Dorea formicigenerans, and Clostridium ramosum. Interestingly, there is no overlap between the “DietPositive” bacteria whose abundance increased after 12 months of a Med Diet reported by Shankar Ghosh et al. [157] and the taxa associated with “healthy diet indices” reported by Asnicar et al. On the other hand, the “DietPositive” and “DietNegative” taxa have substantial overlap with results from our own recent interventional study. We compared the changes in gut microbiome composition after 6 weeks of 500 mg of omega-3 fatty acids or 20 g of soluble fiber (inulin) in 70 middle-age and elderly individuals who had low fiber intake at baseline [158]. We found that, consistent with what is known of prebiotics such as inulin, that Bifidobacterium sp was remarkably increased and this correlated with butyrate production. On the other hand, omega-3 supplementation resulted in a significant decrease in Collinsella sp, both of which are in line with the findings from the multicenter Mediterranean diet intervention [157]. Importantly, many of the changes in bacterial abundances and SCFA concentrations were accompanied by decreases in inflammatory or atherogenic molecular mediators such as VLDL or IL4. Unlike the effects seen through a complex dietary intervention, such as the Mediterranean diet, we showed that it is possible to see significant effects both on microbiome composition and cardiometabolic health mediators.
Other recent studies have highlighted the role of the type of dietary fat on gut microbiome composition. In a randomized trial, 38 overweight and obese subjects were assigned to consume an excess of 1000 kcal/day of diets rich in saturated fat, unsaturated fat, or simple sugars for 3 weeks [159]. The relative abundances of individual taxa were altered in a diet-specific manner, wherein the overfeeding of saturated fat increased Proteobacteria, while unsaturated fat increased butyrate producers.
It is indeed possible to target the increase of specific taxa and the production of specific metabolites and the extent to which different carbohydrate sources (digestible by gut bacteria but not by humans) alter gut microbiome composition and function was recently tested by Deehan et al. [148] in a randomized controlled trial using a four-arm design. They gave increasing doses of three forms of resistant starch from maize, potato, or tapioca and a human-digestible corn starch (placebo) to 10 healthy volunteers per arm. They compared the effect of these carbohydrate substrates on gut microbiome composition in humans and found that chemically modified resistant starches with small structural differences induced highly specific effects on the gut microbiome that direct changes in the output of either propionate or butyrate. Dominant effects were remarkably consistent within treatment groups and dose-dependent with a plateau at a dose of 35 g/day. For example, maize-derived resistant starch resulted in increases in B. adolescentis, E. rectale, Oscillibacter, and Ruminococcus-related taxa and this correlated with increases in butyrate production. Tapioca-resistant starch on the other hand, which induced shifts in propionate proportions, as positively correlated with increases in P. distasonis an important succinate-producing bacterium. Succinate is then converted to propionate by other commensal bacteria in the gut. This very elegant design also clearly demonstrated the dose dependency between carbohydrate sources and the production of specific SCFAs. Similar strong and specific effects on gut microbiome composition have been shown by many interventional studies, both in healthy [160, 161] and diseased populations [162].
Where is this field moving toward?
Over the past decade, considerable evidence from both animal and human studies has accumulated showing a clear link between the gut microbiome in chronic diseases, including inflammatory autoimmune disorders, gut inflammation-related disorders, and cardiometabolic diseases (Fig. 1). It is increasingly clear that bacterial metabolites are at least in part, the key agents involved in the role of the gut microbiome on human health and among such metabolites, SCFAs appear to be the most important ones. Butyrate-producing bacteria are associated with lower risk of inflammatory autoimmune and cardiometabolic diseases, and also irritable bowel syndrome (Table 1). A number of therapeutic strategies to target the gut microbiome are possible, but nutritional changes appear to be the most obvious, noninvasive, and immediate way of altering the gut microbiome composition and function. Recent randomized controlled trials have shown that both composition and function respond in consistent ways to specific dietary interventions. Dietary fiber and unsaturated fat, separately or in a healthy diet such as the Mediterranean diet, result in higher relative abundances of butyrate-producing bacteria and these bacteria and the SCFAs produced in turn result in improved health outcomes. Different types of dietary fiber result in different bacterial changes and different SCFAs. The possibility of designing dietary interventions targeted specifically at increasing certain bacterial metabolites to improve cardiometabolic and inflammatory health outcomes appears well within reach within the next half decade.
People with social anxiety disorder have a unique melange of gut microbes.
If social situations bring on feelings of overwhelming stress, discomfort, and self-consciousness, you’re not alone. Around 15 million adults in the U.S. have social anxiety, a common disorder marked by fear you might be judged or criticized when interacting with other people, whether it’s answering a question in class or shopping at the grocery store.
While social anxiety often pops up during childhood, the exact biological and environmental factors that predispose someone to this condition are still not fully understood. However, some new research suggests the gut microbiome might be involved.
According to a study published this week in the journal Proceedings of the National Academy of Sciences, people with social anxiety disorder have a unique melange of gut microbes that, when transplanted into the guts of mice, resulted in the rodents being more socially fearful than other mice who didn’t get the fecal microbiome transplant.
For their study, researchers at University College Cork in Ireland gathered poop samples from 12 human donors, six individuals had social anxiety disorder, and the other six didn’t. None of the individuals were on any psychiatric medications or supplements that could alter their microbiomes.
Once the researchers did a DNA analysis to confirm that there were considerable microbial differences between the SAD group compared to the control, these samples were transplanted into 72 male mice.
The mice were then put through a ringer of different tests assessing their social behavior and anxiety. The mice with the gut microbes from the group of people with social anxiety disorder didn’t behave any differently than their counterparts with gut microbes from people without the condition. However, there was one test where the two groups differed.
To assess social fear, the researchers shocked the mice with small bursts of electricity when the animals were being introduced to a new mouse. Some days following that interaction, the mice were then introduced to another new mouse. Those animals with the healthy microbiome were unbothered by the conditioning intended with the electric shocks, socializing just fine with their new buddy. The mice with social anxiety disorder microbes, on the other hand, never recovered, remaining fearful of any newcomers.
Delving deeper into the gut-brain connection, the researchers saw that levels of certain hormones that affect the brain, like oxytocin, which promotes social behavior, were lower in the mice with the microbes from those with social anxiety disorder. Genes associated with oxytocin and another hormone involved in social behavior, vasopressin, weren’t as active compared to the mice with healthy donor microbiomes.
The immune system in mice with the microbes from those with social anxiety disorder was also distinct, with changes that seemed to promote inflammation. Previous studies have found that the immune system does play a role in shaping our social behavior.
While this research is still preliminary, it emphasizes how important our gut microbes are to our overall health.
A new antibiotic class that works against multidrug-resistant bacteria has been discovered, offering new opportunities to tackle these dangerous microbes.
The macrocyclic peptide zosurabalpin showed promising antibacterial activity against Carbapenem-resistant Acinetobacterbaumannii (CRAB), according to two papers published in the journal Nature.
The Gram-negative bacteria is deemed an urgent threat by the U.S. Centers for Disease Control and Prevention and has also been classified as a priority 1 critical pathogen by the World Health Organization.
“The new molecule overcomes the existing drug-resistance mechanisms that the currently available antibiotics are failing to address,” said Kenneth Bradley, global head of infectious disease discovery at Roche pharma research & early development, which worked with Harvard researchers to establish the mechanism of action for the compound.
“With this significant breakthrough, zosurabalpin has the potential to address a major unmet need in the fight against antimicrobial resistance.”
Gram-negative bacteria are particularly difficult to kill because they are encased in both inner and outer membranes that are challenging for most antibiotics to cross. The cytoplasmic membrane is surrounded by an outer membrane containing lipopolysaccharide (LPS), which blocks the entry of most antibiotics.
Zosurabalpin traps this LPS during its transport to the outer membrane by inhibiting a complex of proteins called the LptB2FGC complex.
Specifically, the clinical candidate binds to both LPS and a transport complex that facilitates movement to its destination. The LPS transport complex is trapped in a substrate-bound state, which leaves it unable to move LPS and results in death of the bacterium.
A team led by Claudia Zampaloni, PhD, senior principal scientist in infectious diseases at Roche, initially examined around 45,000 tethered macrocyclic peptides, which have greater molecular weights than most antibiotics.
The researchers identified one that selectively killed A. baumannii, which was further honed for efficacy and tolerability using a new type of test based on blood-plasma compatibility, culminating in the discovery of zosurabalpin.
The drug was effective against more than 100 CRAB clinical samples tested in the lab and in multiple mouse infection models, including sepsis and thigh and lung infection induced by CRAB strains.
In another research article, Karanbir Pahil, PhD, from Harvard University, and colleagues use X-ray techniques to identify structures that show zosurabalpin engages LptB2FGC only when the complex is bound to LPS, indicating the importance of the latter for its action.
The basis for its specificity against A. baumannii lies in this protein complex, explaining why it is ineffective against other bacteria whose Lpt proteins may have different amino-acid sequences.
This previously unknown mode of antibiotic action suggests that pre-existing resistance is unlikely, note Morgan Gugger and Paul Hergenrother, PhD, both from the University of Illinois at Urbana-Champaign, in accompanying News and Views article.
They point out that the U.S. Food and Drug Administration has not approved any new classes of antibiotic for harmful Gram-negative bacteria in more than 50 years.
“Treatment options for CRAB infections continue to dwindle as mortality rates are rising, with some estimated death rates reaching approximately 50% for invasive infections,” they warn.
The commentators add that zosurabalpin might not have the damaging effect on normal gut microbes seen with most antibiotics, given its high specificity for A. baumannii.
“The movement towards bacterium-specific antibiotics is a new development, and one that can be facilitated by diagnostics that can rapidly identify specific harmful bacteria in infected individuals,” they wrote.
“Given that zosurabalpin is already being tested in clinical trials, the future looks promising, with the possibility of a new antibiotic class being finally on the horizon for invasive CRAB infections.”







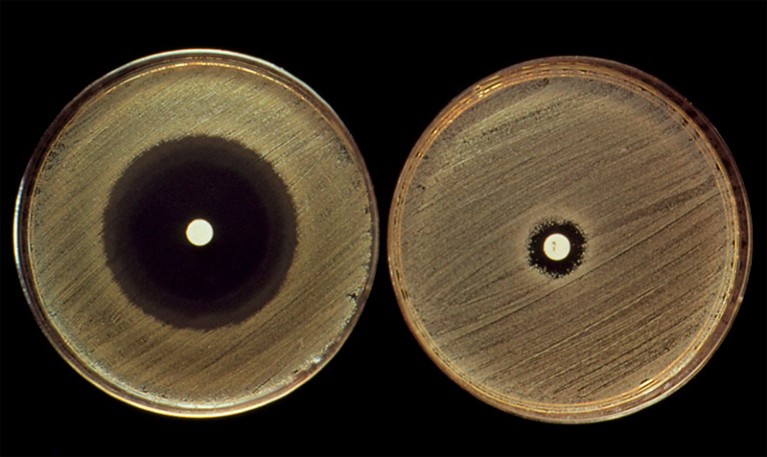


 By Robin Foster HealthDay Reporter
By Robin Foster HealthDay Reporter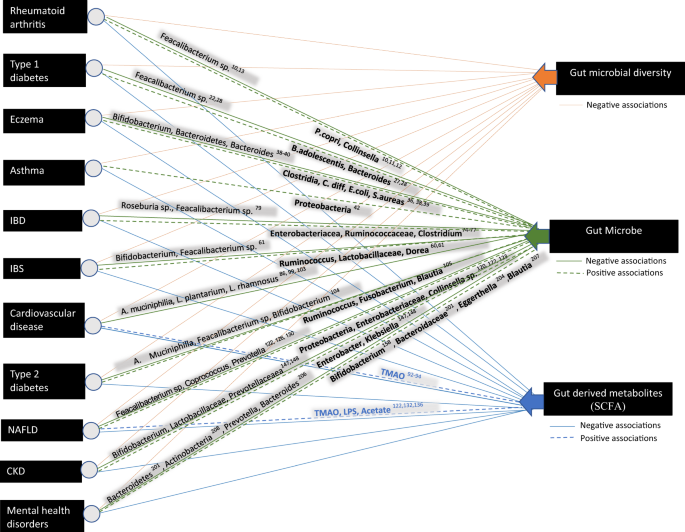



{kind=link}
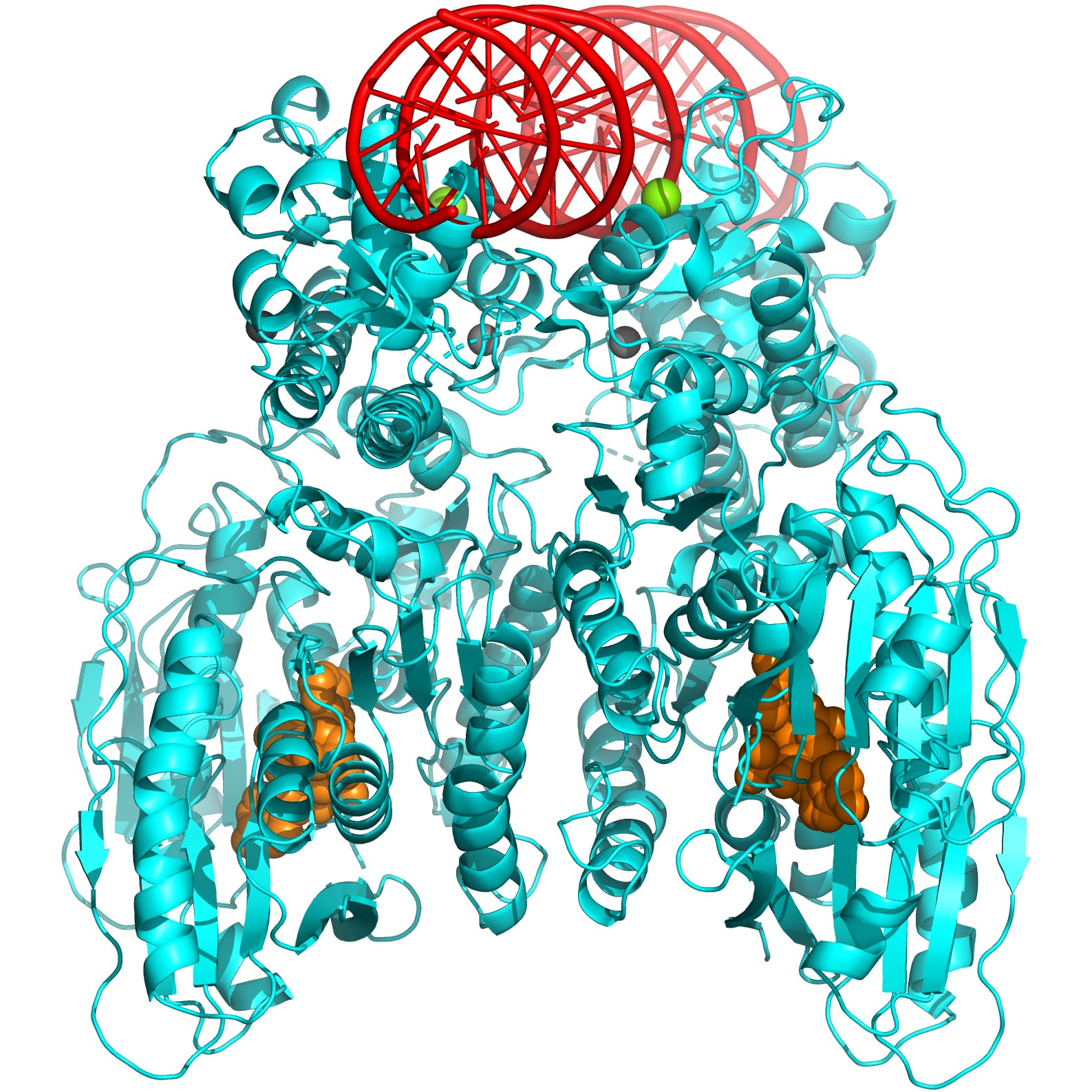{kind=link}
{kind=link}