The adult mammalian brain can produce new neurons in a process called adult neurogenesis, which occurs mainly in the subventricular zone (SVZ) and in the hippocampal dentate gyrus (DG). Brain-derived neurotrophic factor (BDNF) signaling and cannabinoid type 1 and 2 receptors (CB1R and CB2R) have been shown to independently modulate neurogenesis, but how they may interact is unknown. We now used SVZ and DG neurosphere cultures from early (P1-3) postnatal rats to study the CB1R and CB2R crosstalk with BDNF in modulating neurogenesis. BDNF promoted an increase in SVZ and DG stemness and cell proliferation, an effect blocked by a CB2R selective antagonist. CB2R selective activation promoted an increase in DG multipotency, which was inhibited by the presence of a BDNF scavenger. CB1R activation induced an increase in SVZ and DG cell proliferation, being both effects dependent on BDNF. Furthermore, SVZ and DG neuronal differentiation was facilitated by CB1R and/or CB2R activation and this effect was blocked by sequestering endogenous BDNF. Conversely, BDNF promoted neuronal differentiation, an effect abrogated in SVZ cells by CB1R or CB2R blockade while in DG cells was inhibited by CB2R blockade. We conclude that endogenous BDNF is crucial for the cannabinoid-mediated effects on SVZ and DG neurogenesis. On the other hand, cannabinoid receptor signaling is also determinant for BDNF actions upon neurogenesis. These findings provide support for an interaction between BDNF and endocannabinoid signaling to control neurogenesis at distinct levels, further contributing to highlight novel mechanisms in the emerging field of brain repair.
Introduction
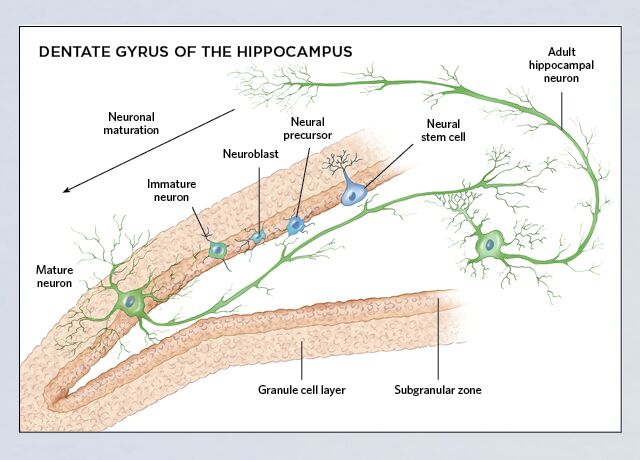
Constitutive neurogenesis occurs in the adult mammalian brain where NSPC are able to differentiate into three neural lineages, neurons, astrocytes and oligodendrocytes (Gage, 2000; Gross, 2000). These multipotent cells exhibit properties of self-renewal and cell proliferation that allow the maintenance of their own pool (Ma et al., 2009). Neurogenesis occurs mainly in two brain areas, the subventricular zone (SVZ) and the subgranular zone (SGZ) within the DG of the hippocampus. These regions are packed with NSPC that originate neuroblasts which migrate toward their final destinations, where they differentiate into mature neurons and are integrated into the neuronal circuitry (Lledo et al., 2006; Zhao et al., 2008; Ming and Song, 2011).
Adult neurogenesis and the neurogenic niches are highly regulated by several factors (intrinsic and extrinsic factors) that control the NSPC rates of proliferation, lineage differentiation, migration, maturation and survival (Ming and Song, 2011). Knowing and understanding the actions of these factors will further contribute to develop new therapeutic strategies useful for brain repair and regeneration. However, there is still a lack of knowledge regarding the key factors that regulate each step of postnatal neurogenesis.
The role of neurotrophins and, in particular, brain-derived neurotrophic factor (BDNF) in adult neurogenesis has been the subject of many studies (Henry et al., 2007; Chan et al., 2008; Vilar and Mira, 2016). BDNF is expressed in both SVZ and SGZ neurogenic niches (Galvão et al., 2008; Li et al., 2008) but its precise role in adult neurogenesis is still not consensual. In fact, some studies suggest that BDNF is important to positively regulate DG cell proliferation and survival (Chan et al., 2008; Li et al., 2008) while others report no BDNF-induced changes in DG neurogenesis (Choi et al., 2009). In SVZ, most studies depict that BDNF does not promote any significant changes in cell proliferation and survival (Henry et al., 2007; Galvão et al., 2008), despite having a role in the migration of SVZ-derived cells (Snapyan et al., 2009; Bagley and Belluscio, 2010). Despite the available contradictory data, BDNF, through TrkB signaling, was shown to have an essential role in the regulation of dendritic complexity as well as synaptic formation, maturation and plasticity of newborn neurons (Chan et al., 2008; Gao et al., 2009; Wang et al., 2015).
Besides expressing BDNF, NSPC present in the neurogenic niches were shown to express all the elements of the endocannabinoid system (Aguado et al., 2005; Arévalo-Martín et al., 2007), including the main cannabinoid receptors type 1 (CB1R) and type 2 (CB2R) receptors (Rodrigues et al., 2017). They are both present in the CNS, although CB2R expression is relatively higher in the immune system (Galve-Roperh et al., 2007). In recent years, the role of cannabinoids in neurogenesis has been of particular interest given their multiplicity of neuromodulatory functions (Mechoulam and Parker, 2013). Cannabinoid receptors modulate adult neurogenesis by acting at distinct neurogenic phases (Prenderville et al., 2015). Importantly, activation of type 1 (Xapelli et al., 2013) or type 2 cannabinoid receptors (Palazuelos et al., 2006) by selective agonists was found to regulate cell proliferation, neuronal differentiation and maturation (Rodrigues et al., 2017).
Several studies have provided molecular and functional evidence for a crosstalk between BDNF and endocannabinoid signaling (Maison et al., 2009; Zhao et al., 2015). Synergism between BDNF and CB1R has been observed both in vitro and in vivo (De Chiara et al., 2010; Galve-Roperh et al., 2013). In particular, BDNF was shown to regulate striatal CB1R actions (De Chiara et al., 2010). Moreover, evidence for BDNF-TrkB signaling interplay with CB1R has been shown to trigger endocannabinoid release at cortical excitatory synapses (Yeh et al., 2017). Importantly, genetic deletion of CB1R was shown to promote a decrease in BDNF expression (Aso et al., 2008) while induction of BDNF expression contributed to the protective effect of CB1R activity against excitotoxicity (Marsicano, 2003; Khaspekov et al., 2004). Moreover, CB1R activity can enhance TrkB signaling partly by activating MAP kinase/ERK kinase pathways (Derkinderen et al., 2003) but also by directly transactivating the TrkB receptors (Berghuis et al., 2005). Δ9-THC, the principal active component of cannabis, was shown to promote upregulation of BDNF expression (Butovsky et al., 2005) whereas increased levels of BDNF were shown to rescue the cognitive deficits promoted by Δ9-THC administration (Segal-Gavish et al., 2017). Interestingly, clinical data suggests that acute and chronic intermittent exposure to Δ9-THC alters BDNF serum levels in humans (D’Souza et al., 2009).
Given the evidence that BDNF and cannabinoid signaling can affect neurogenesis as well as the fact that BDNF may interact with cannabinoid receptors, we hypothesized that cannabinoid receptors could act together with BDNF signaling to fine-tune neurogenesis. We show for the first time that endogenous BDNF is crucial for the cannabinoid-mediated effects on SVZ and DG neurogenesis to happen. Moreover, we demonstrate that CB2R has a preponderant role in regulating some of the BDNF actions on neurogenesis. Taken together, our results suggest an important crosstalk between BDNF and cannabinoid signaling to modulate postnatal neurogenesis.
Discussion
The present work reveals a yet not described interaction between BDNF and cannabinoid receptors (CB1R and CB2R) responsible to modulate several aspects of SVZ and DG postnatal neurogenesis. BDNF was shown to be an important modulator of SVZ and DG postnatal neurogenesis, its actions being under control of cannabinoid receptors. The relevance of each cannabinoid receptor to control the action of BDNF upon neurogenesis is different in the two neurogenic niches. While CB2R has a preponderant role in modulating BDNF actions on DG, BDNF-mediated SVZ postnatal neurogenesis is modulated by both CB1R and CB2R. A constant and clear finding in both neurogenic niches is that BDNF is required for cannabinoid actions to occur. It thus appears that a reciprocal cross-talk between cannabinoids and BDNF exist to modulate postnatal neurogenesis.
BDNF is a neurotrophin important in the regulation of several neuronal processes such as neuronal branching, dendrite formation and synaptic plasticity (Dijkhuizen and Ghosh, 2005; Gómez-Palacio-Schjetnan and Escobar, 2013). In line with this evidence, several studies have shed light on the actions of BDNF in the survival and differentiation of newborn neurons (Benraiss et al., 2001; Henry et al., 2007; Chan et al., 2008; Snapyan et al., 2009). Our findings now demonstrate that BDNF is able to affect early steps of postnatal neurogenesis, such as cell-fate, cell proliferation and neuronal differentiation of SVZ and DG cultures. We observed that BDNF promoted self-renewal of SVZ- and DG-derived cells as observed by an increase in self-renewal divisions, i.e., an increase in the percentage of Sox2+/+ cell-pairs. BDNF-CBR crosstalk has been reported to control several processes at the synaptic level (Zhao and Levine, 2014; Zhong et al., 2015) and we now extended these findings toward very early stages of postnatal neurogenesis. Interestingly, the increase in the SVZ and DG pool of stem/progenitor cells mediated by BDNF was fully abolished in the presence of CB2R antagonist but not CB1R antagonist. An exception is the influence of BDNF upon SVZ cell proliferation, which is not affected by CB1R or CB2R selective antagonism. In what concerns neuronal differentiation, both CB1R and CB2R are required for BDNF actions on SVZ whereas at the DG, only CB2R seem to affect BDNF-promoted neuronal differentiation. Overall, cannabinoid receptor blockade appears to influence more BDNF-induced actions upon early stages of DG neurogenesis in comparison to SVZ, highlighting the fact that cannabinoids distinctly modulate the effects promoted by BDNF in SVZ and DG neurogenesis.
It was previously known that the endocannabinoid system and cannabinoid receptors are important modulators of several stages of neurogenesis (Palazuelos et al., 2012; Xapelli et al., 2013; Prenderville et al., 2015; Rodrigues et al., 2017). In accordance with our previous data, SVZ and DG cells were differently affected by the same cannabinoid pharmacological treatments (Rodrigues et al., 2017). Considering cell fate, we observed that selective activation of CB2R activation promotes self-renewal of DG cells, but not of SVZ cells. This is consistent with several pieces of evidence showing a regulation of cell fate promoted by the activation of several signaling pathways [such as mitogen-activated protein kinase (MAPK) family (ERK, JNK and p38) and the phosphoinositide-3 kinase (PI3K)/AKT pathways] triggered by CBR activation (Molina-Holgado et al., 2007; Gomez et al., 2010; Soltys et al., 2010; Compagnucci et al., 2013).
On the other hand, our results reveal, for the first time, a role of cannabinoid receptors (CB1R and CB2R) in regulating DG cell commitment.
Considering cell proliferation, it is promoted by CB1R but not CB2 at SVZ, while at DG cell proliferation was only induced by co-activation of CB1R and CB2R. These results are in accordance with previous reports that have shown an increase in SVZ cell proliferation promoted by CB1R selective activation (Trazzi et al., 2010; Xapelli et al., 2013) and an increase in DG cell proliferation triggered by CB1R/CB2R non-selective activation (Aguado et al., 2005; Rodrigues et al., 2017). Importantly, while we also detected an effect with the non-selective CB1R/CB2R agonists, none of the selective agonists when applied in the absence of the other agonist were effective to promote cell proliferation in the DG, highlighting the need of caution while interpreting negative results with each of those agonists separately.
Regarding neuronal differentiation, our data indicate that in SVZ and DG neurogenic niches both subtypes of cannabinoid receptors are able to promote neuronal differentiation. These data are in accordance with previous reports in which cannabinoid receptor activation enhanced neuronal differentiation of NSPC by CB1R- (Compagnucci et al., 2013) or CB2R-dependent (Avraham et al., 2014) mechanisms.
The most important finding in the present work is that most of the cannabinoid-induced effects upon cell proliferation and neuronal differentiation depend on the presence of BDNF, suggesting the existence of a BDNF-endocannabinoid feedback loop responsible for regulating these processes. Previous reports have shed light on the existence of a putative interaction between BDNF and cannabinoid receptors (Howlett et al., 2010), but none focused upon neurogenesis. De Chiara et al. (2010) have identified a novel mechanism by which BDNF mediates the regulation of striatal CB1R function. Moreover, others have suggested that BDNF can regulate neuronal sensitivity to endocannabinoids through a positive feedback loop important for the regulation of neuronal survival (Maison et al., 2009). Evidence also shows the involvement of BDNF in the actions mediated by cannabinoids against excitotoxicity (Khaspekov et al., 2004), in synaptic transmission and plasticity (Klug and van den Buuse, 2013; Zhao et al., 2015; Yeh et al., 2017) and in several behavioral outputs (Aso et al., 2008; Bennett et al., 2017). Previous animal studies have shown that acute (Derkinderen et al., 2003) and chronic (Butovsky et al., 2005) Δ9-THC (major psychoactive constituent of cannabis; CB1R and CB2R agonist) administration is associated with an increase in BDNF gene expression. Moreover, it was shown that overexpression of BDNF is able to rescue cognitive deficits promoted by Δ9-THC administration in a mouse model of schizophrenia (Segal-Gavish et al., 2017). In human studies it was found that Δ9-THC increased serum BDNF levels in healthy controls, but not in chronic cannabis users (D’Souza et al., 2009). In fact, cyclic AMP response element-binding protein (CREB) may be the common linking element because it is an important regulator of BDNF-induced gene expression (Finkbeiner et al., 1997), and has been reported to control several steps of the neurogenic process in the adult hippocampus (Nakagawa et al., 2002) and SVZ (Giachino et al., 2005). Consistently, cannabinoids have been shown to induce CREB phosphorylation (Isokawa, 2009) and also to promote changes in BDNF and CREB gene expression (Grigorenko et al., 2002). In addition, the work done by Berghuis et al. (2005) showed that endocannabinoids stimulate TrkB receptor phosphorylation during interneuron morphogenesis. Most importantly, in the same study, the authors observed by co-immunoprecipitation the formation of heteromeric complexes in PC12 cells expressing TrkB receptors and CB1R (Berghuis et al., 2005). Our study brings new and relevant information on the interaction between cannabinoid receptors and BDNF in controlling SVZ and DG neurogenesis, and clearly highlights that this interaction is reciprocal. In fact, neurogenesis promoted by cannabinoid receptor activation depends on the presence of endogenous BDNF, while the effects mediated by BDNF upon neurogenesis are directly regulated by modulation of CB1R or CB2R.
Although our study is based on an in vitro approach, the neurosphere assay, it represents a highly relevant model. In vitro systems of NSPC allow an easier access and better control of experimental variables as well as a thorough analysis of mechanisms happening at cellular and molecular level providing useful information to be further validated in vivo (Singec et al., 2006). Moreover, the heterogeneous composition of the NSPC grown in neurospheres is extremely relevant because it holds some of the features, such as close contact with neighboring cells (newly generated neuroblasts, astrocytes and oligodendrocytes), that resemble those of the physiological niche (Casarosa et al., 2014). These well-established advantages (Aguado et al., 2007; Agasse et al., 2008; Azari et al., 2010) are the reason why we have used this in vitro approach to study the intrinsic properties of NSPC and to understand the interaction between BDNF and cannabinoids in modulating neurogenesis. It is, however, important to mention that the mechanisms governing the regulation of neurosphere dynamics might be different from the ones regulating in vivo adult neurogenesis (Casarosa et al., 2014). Indeed, further in vivo studies will be required to comprehensively understand the role of BDNF in regulating the actions of cannabinoid receptors on postnatal neurogenesis.
Taken together, our data highlight a novel level of complexity for the regulatory mechanisms involved in NSPC dynamics, which involve the interplay of multiple signaling cues, and where BDNF and cannabinoids may play a relevant role. Further in vitro studies are required to detail the molecular mechanisms involved, as well as in vivo studies to determine the functional consequences of the BDNF/cannabinoid crosstalk to control neurogenesis. Nevertheless, our study provides evidence for the need of integrative strategies whenever focusing on NSPC for brain repair.